PART 1
*by Maren Wellenreuther, Anna Runemark and Mikkel Brydegaard
We study a
variety of species in our lab, such as ecotypes of isopods and color morphs of
lizards and damselflies. In our work on these species, color is one of the main
traits that we examine. This is because color affects crypsis (isopods), sexual
selection and other life history traits (lizards and damselflies). However,
examining color is not a trivial undertaking. For example, biologists often
fail to account for differences in animal visual systems when modeling how
species are perceived by con- and heterospecifics. Many also fail to realize
that most cameras are not sufficiently spectrally
resolved (they have 3 bands only) to capture color and that most
spectrophotometers are not sufficiently spatially
resolved to capture heterogeneity in color patterns (since they give averages
rather than spatially resolved measures, consequently, both a donkey and a
zebra would appear grey to a high resolution spectrometer).

Ecotypes of isopods Asellus aquaticus, color morphs of the Skyros Wall Lizard Podarcis gaigeae, and different colors a
mating couple of the structurally colored Calopteryx
virgo.
The beginnings…
When we
first started to measure color, we realized how difficult this task actually
is. This is particular true when one wants to conduct color experiments in
natural or laboratory settings. What light sources are needed? What color
vision does the color-signal-receiver have? Faced with these problems, we
quickly recognized that we needed to communicate with people that know how to measure color. The Atomic
Physics Section at Lund
University has a group
(Applied molecular spectroscopy and remote sensing) under the supervion of Sune
Svanberg that, among other things, specializes in the measurement of color.
First contact with the group was established in 2007 when Fabrice Eroukhmanoff
was hoping to quantify the color of isopod ecotypes, of which one ecotype
typically inhabits the reed habitat, while the other one prefers chara habitat.
Fabrice wanted to quantify the color of different ecotypes. After talking to Mikkel
Brydegaard Sørensen, a PhD student in Sune’s group, they realized that in order
to compare the photographs, he needed to standardize them posteriori to account for different light environments. The color
analyses are presented in, for example, here.
Discussions and exchange of ideas between our group and Sune’s group helped in
the years to come to set up carefully planned experiments to examine color
traits.
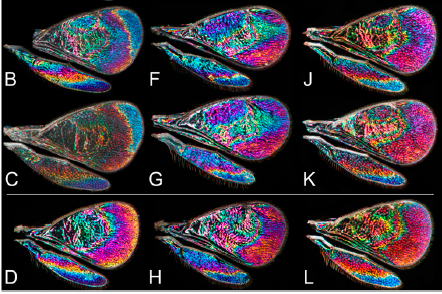
To illustrate the point…measuring complex color traits in the Skyros Wall lizard
For Anna
Runemark, who is studying color morphs of the Skyros Wall lizard but also
dorsal coloration, color is one of the core traits of her studies. In her work,
she is interested in the strength and direction of sexual selection on
different color morphs in island and mainland populations (see here).
To measure color differences, Mikkel designed an optically isolated ‘photo-box’
with a standardized light environment (e.g. the only illumination source is the
flash) and polarizing filters in front of the flash and the objective to avoid
specular reflectance. As the distances from the flash vary with position in the
box, the white background is used to interpolate an illumination profile on the
lizard, and this was used to correct for illumination differences across the
photograph. Lastly, to estimate the entire color probability distribution
(includes information about both mean, variance, skewness, kurtosis etc., see
e.g. the 2D examples in the figure below), the color of each pixel of the
measured colour patch was quantified, spatially accumulated and then divided by
the number of pixels to obtain the probability. This method is presented in a
paper found here

Figure 4-taken from (Brydegaard, Runemark et al. 2012). Upper row: reflectance distributions for a
homogeneously colored yellow-throated lizard. Lower row: corresponding
distributions for a orange-yellow patchy specimen. Left column: three 1D
distributions for each spectral band. Middle column: 2D chromatic plane
distributions color coded with corresponding colors for the two example
specimen. Right column: iso-surfaces encapsulating probabilities higher than 1%
in the 3D RGB color space of the two sample specimens. Surfaces are coded with
corresponding colors.
After these
initial dialogs between biologists and physicist, it became clear that we can
both learn from each other. We as biologists have an interest in capturing
color traits of animals accurately and precisely, while physicist that study
natural phenomena have a need to understand the biological underpinnings. As a
result of this initial integrative work, the Lund CAnMove group organized a
symposium entitled ”The Biology-Physics Interface”. During this day, many of us
met and presented our research and discussed ideas.


Caption: CAnMove symposium ”The Biology-Physics
Interface”. Sune Svanberg at the top, and Erik Svensson below.
References
Brydegaard, M., A. Runemark, et al. (2012). "Chemometric approach to chromatic spatial variance. Case study: patchiness of the Skyros wall lizard." Journal of Chemometrics
Eroukhmanoff, F., A. Hargeby, et al. (2009). "Parallelism and historical contingency during rapid ecotype divergence in an isopod." Journal of Evolutionary Biology 22(5): 1098-1110.
Runemark, A. and E. I. Svensson (2012). "Sexual selection as a promoter of population divergence in male phenotypic characters: a study on mainland and islet lizard populations." Biological Journal of the Linnean Society 106(2): 374-389.